It has long been known that there is very long-term cyclical behaviour in a variety of geological records, including mass extinction events, stratigraphic sequences, and climate change. The lengths of these inferred cycles are approximately 60 million years, and 140 million years. Raup and Sepkoski argued for a cycle length of about 28 million years for major extinction events, and proposed the "Death Star" hypothesis, in which a companion star in mutual orbit with the Sun would cause catastrophic meteorite showers resulting in these extinctions.
The Death Star hypothesis has apparently been disproved--at least the Sun's companion has not been discovered despite prudent searching.
In the recent issue of Eos (only available by paid subscription, but your university library probably has it) the headline article suggests that the long-term cyclicity noted above may be present in mantle upwelling events.
The method of analysis makes much use of wavelet analysis, which reminiscent of Fourier analysis but also takes into consideration the portions of the time series in which the periodicities occur.

In the figure above, the lighter colours (green to red) represent significant peaks in cyclicity in a) number of marine genera (caused mainly by episodes of sudden loss and more gradual replacement) and b) the timing of the onset of large igneous provinces (which means eruption of huge volumes of magma). The period of the observed cycles is on the scale at left. The method allows us to consider that the cycles may only be active over portions of the records.
As you get closer to the ends of the record, there is a real possibility that the truncation of your data set will affect the data. The longer the period under investigation, the greater the impact on the data near the beginning and end of the record. Consequently, the yellow dashed arcs in both figures above show us the limit of viable interpretation--we only take seriously the portion of the graph in the "bowl" of the yellow arc.
Although the authors do not describe a precise mechanism by which such cyclicity may arise, they note that previous studies suggest that instabilities in the lowest level of the mantle may occur in a repetitive fashion, leading to cyclicity in the emplacement of large igneous provinces (eruptions that may cover > 100,000 sq-km in a geologically short interval).
Alternatively, the formation of supercontinents can act as a cap on the heat produced within the earth, resulting in mantle upwelling within the continent leading to emplacement of huge volumes of magma and rifting. The supercontinent cycle lasts from 300 to 500 million years and is thus too long to be recognizable on the above graphs.
The sudden appearance of cyclicity may be an example of an emergent property. As previously discussed, emergent properties are global-scale behaviours which arise in a manner which is not at all understood from the interaction of local subsystems. Interestingly, small changes at the local scale may bring about structural change at the global scale, which I have previously referred to as innovation.
Kirchner, J. W., 2002. Evolutionary speed limits inferred from the fossil record. Nature, 415: 65-68.
Rampino, M. R. and Prokoph, A., 2013. Are mantle plumes periodic? Eos, Transactions of the American Geophysical Union, 94 (12): 113-114.
The Death Star hypothesis has apparently been disproved--at least the Sun's companion has not been discovered despite prudent searching.
In the recent issue of Eos (only available by paid subscription, but your university library probably has it) the headline article suggests that the long-term cyclicity noted above may be present in mantle upwelling events.
The method of analysis makes much use of wavelet analysis, which reminiscent of Fourier analysis but also takes into consideration the portions of the time series in which the periodicities occur.

Wavelet analysis of biological diversity (top) compared with that
of initiation of large igneous provinces (i.e., really big eruptions)
over the last 525 million years. From Rampino and Prokoph (2013).
In the figure above, the lighter colours (green to red) represent significant peaks in cyclicity in a) number of marine genera (caused mainly by episodes of sudden loss and more gradual replacement) and b) the timing of the onset of large igneous provinces (which means eruption of huge volumes of magma). The period of the observed cycles is on the scale at left. The method allows us to consider that the cycles may only be active over portions of the records.
As you get closer to the ends of the record, there is a real possibility that the truncation of your data set will affect the data. The longer the period under investigation, the greater the impact on the data near the beginning and end of the record. Consequently, the yellow dashed arcs in both figures above show us the limit of viable interpretation--we only take seriously the portion of the graph in the "bowl" of the yellow arc.
Although the authors do not describe a precise mechanism by which such cyclicity may arise, they note that previous studies suggest that instabilities in the lowest level of the mantle may occur in a repetitive fashion, leading to cyclicity in the emplacement of large igneous provinces (eruptions that may cover > 100,000 sq-km in a geologically short interval).
Alternatively, the formation of supercontinents can act as a cap on the heat produced within the earth, resulting in mantle upwelling within the continent leading to emplacement of huge volumes of magma and rifting. The supercontinent cycle lasts from 300 to 500 million years and is thus too long to be recognizable on the above graphs.
Both graphs above suggest the sudden appearance of an oscillation with a period on the order of 30-35 My beginning a little less than 150 million years ago. That kind of sudden appearance of a new phenomenon in the geological record is an example of innovation in the earth system.

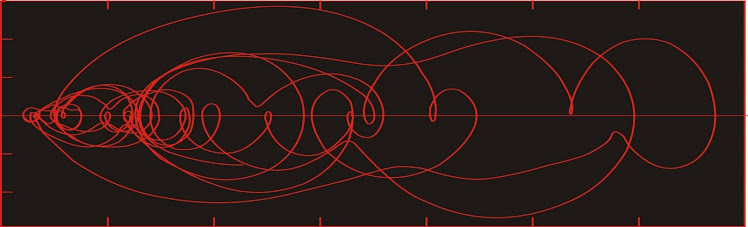
In an earlier post (based on the variograms produced by Kirchner, 2002), I argued that extinction was more-or less random, on the basis of the lack of a consistent slope. There were some spectral peaks which could be correlated to the cycles described above--the ca. 28-million-year cyclicity is probably related to the 30-million-year periodicity noted above. An additional oscillation with a period of about 140 million years is also present.
Plot of spectral power of extinctions vs originations. After Kirchner (2002).
The sudden appearance of cyclicity may be an example of an emergent property. As previously discussed, emergent properties are global-scale behaviours which arise in a manner which is not at all understood from the interaction of local subsystems. Interestingly, small changes at the local scale may bring about structural change at the global scale, which I have previously referred to as innovation.
Kirchner, J. W., 2002. Evolutionary speed limits inferred from the fossil record. Nature, 415: 65-68.
Rampino, M. R. and Prokoph, A., 2013. Are mantle plumes periodic? Eos, Transactions of the American Geophysical Union, 94 (12): 113-114.
